- Peak performance
- The science
- Healthcare
HomePeak performanceThe scienceStoriesAffiliate programAboutNewsEventsPartnersJobsSupportSign in
Peak performance
Heathcare
The science
Prepare to Timeshift®


Light is perhaps the single most powerful force shaping human biology. Long before alarm clocks, shift patterns, or smartphones existed, the daily rise and fall of sunlight orchestrated virtually every aspect of our physiology — from when we sleep and wake, to when our hearts beat fastest, our hormones surge, and our bodies digest food. That ancient partnership between light and life is mediated by an exquisitely tuned timekeeping system known as the circadian clock. Understanding how light interacts with this clock is one of the most important questions in modern sleep and chronobiology research, with profound implications for health, medicine, and society.
This article reviews the science of light and circadian rhythms in humans, and explains what circadian rhythms are, how the eye detected light for timekeeping long before the evolution of vision, what properties of light matter most, and what happens when the timing of the light-dark cycle goes wrong — whether due to blindness, transatlantic flights, night shifts, spaceflight, or urban light pollution.
Circadian rhythms – rhythms with a cycle length of approximately 24 hours - are defined as self-sustaining biological oscillations that persist even in the complete absence of external time cues, i.e., they are internally generated spontaneously and do not need any outside stimulus to drive the rhythm [1]. They are found in virtually every living organism — from single-celled algae to humans — and represent one of the most conserved features of life on Earth [2].
In humans, the central circadian clock resides in a tiny bilateral structure in the hypothalamus called the suprachiasmatic nucleus (SCN), a cluster of approximately 50,000 neurons located just above the optic chiasm [3]. Each SCN neuron functions as an independent oscillator, driven by interlocking feedback loops of multiple "clock genes" that cycle with an intrinsic period close to, but rarely exactly, 24 hours [4]. The average intrinsic period across the human population is approximately 24.2 hours, ranging from about 23.5 to 25.0 hours [5]. As the period of the internal clock is not exactly 24 hours, it must be reset, or ‘entrained’, every day by external time cues, the most powerful of which is the daily light-dark cycle.
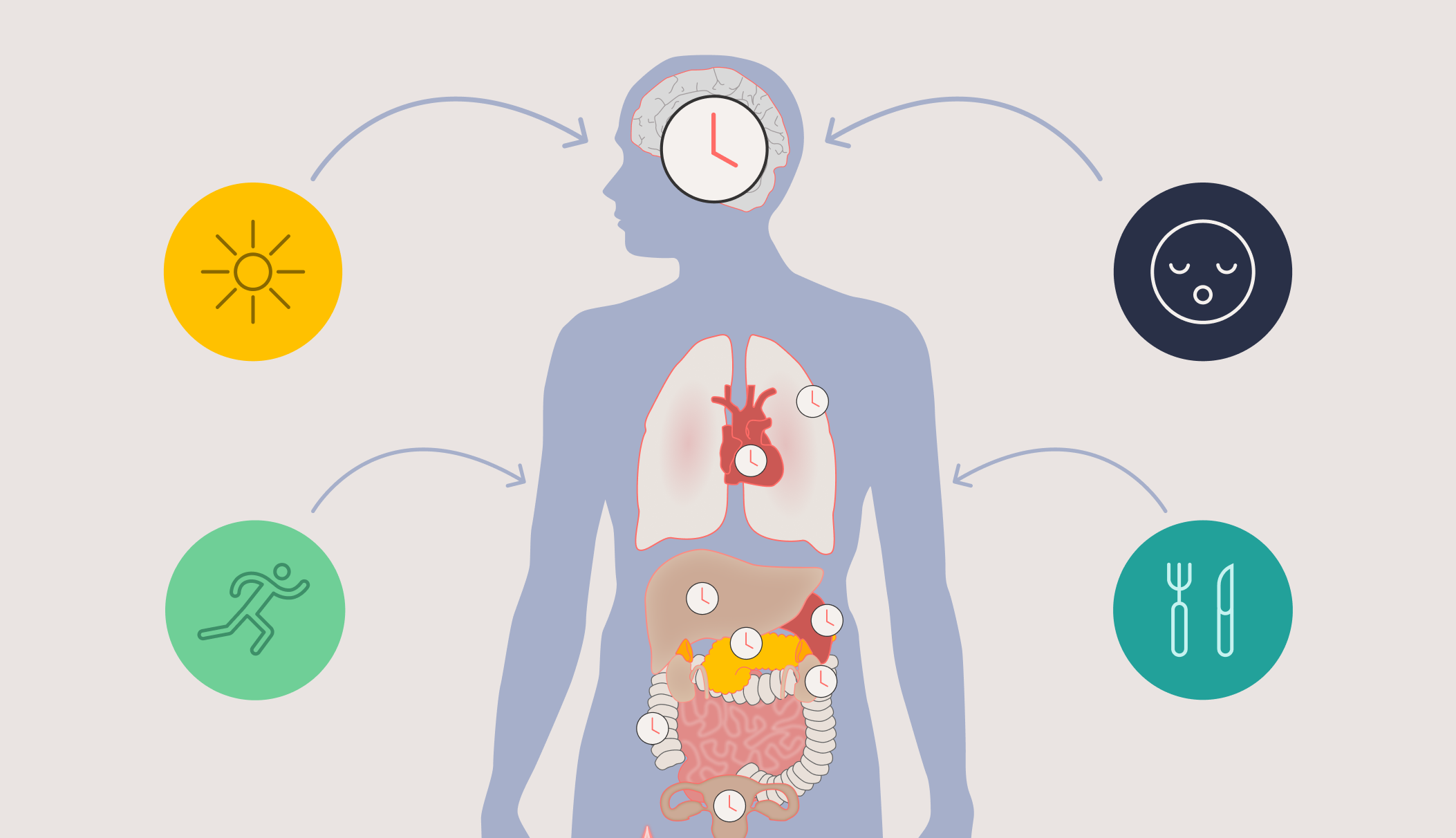
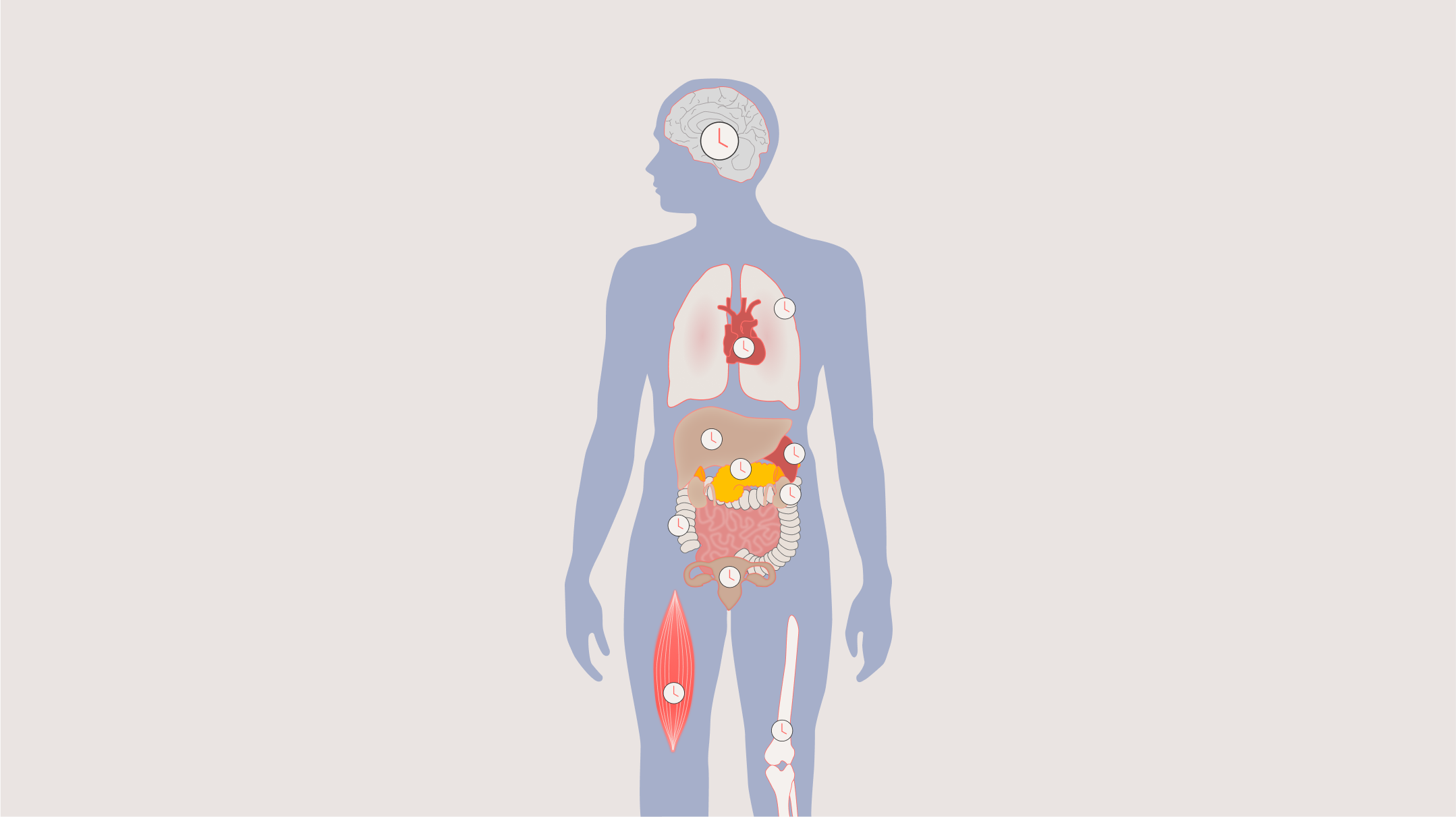
The SCN generates signals that synchronise many physiological and behavioural rhythms including sleep, alertness, performance and mood; core body temperature; many hormone rhythms; glucose and lipid regulation; immune responses and even reproductive function [6].
Virtually every tissue and organ also contains additional peripheral circadian clocks that help keep local time, for example in the heart, lungs, liver and skin, and the SCN acts as a central conductor, keeping all the different rhythms across the brain and body in time together. The rhythms that result are not trivial background fluctuations; they represent fundamental temporal organisation of our entire biology.
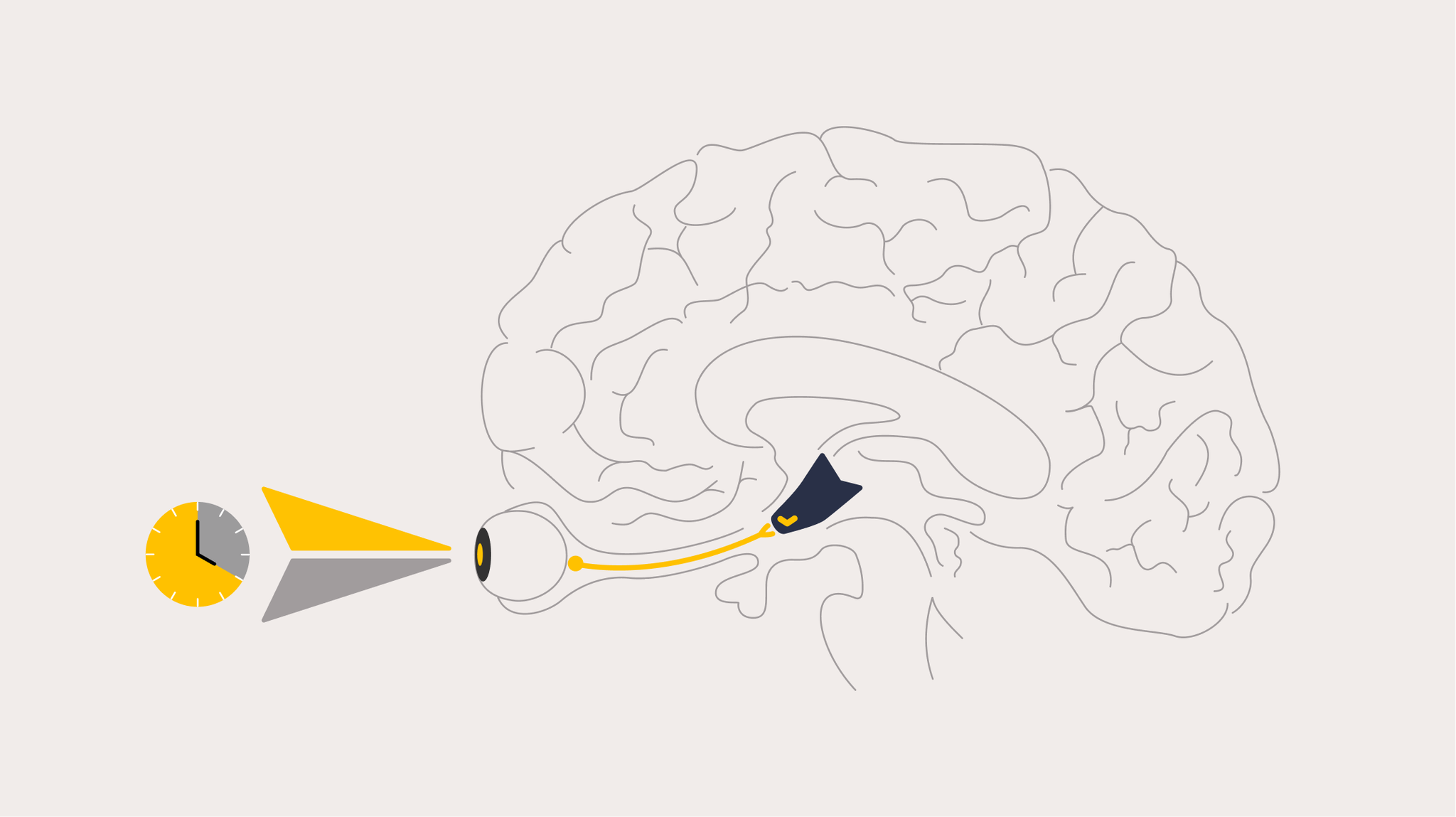
Light information reaches the SCN directly from the eyes via a dedicated neural pathway called the retinohypothalamic tract (RHT), which projects directly from specialized photoreceptor cells in the retina to the SCN [7]. This pathway is anatomically and functionally separate from the visual pathway that generates our conscious perception of images, and it was not fully characterised until around 25 years ago.
Critically, total blindness, whether through bilateral enucleation or damage to the eye that prevents any eye function, abolishes the ability of light to reset the clock, including in humans, confirming that only the eyes can detect light for circadian entrainment and other non-visual effects [8].
The existence of this dedicated non-visual light pathway was powerfully demonstrated by studies showing that circadian responses to light could survive complete loss of rod and cone photoreceptors, the ones we use to see [9-13]. There are a small number of ‘totally’ blind individuals who retain circadian responses to light despite not having any conscious light perception or any measurable rod or cone function. When the first such case was reported [9], scientists did not understand how the light was being detected by the eye. The search for the mystery photoreceptor mediating this response ultimately led to the discovery of intrinsically photosensitive retinal ganglion cells (ipRGCs).
Around the turn of the millennium, a small subpopulation of retinal ganglion cells was found to contain a photopigment called melanopsin [14] which makes them directly light-sensitive, unlike the vast majority of ganglion cells, which only respond to light indirectly via signals from rods and cones [15]. The cells are called intrinsically photosensitive retinal ganglion cells (ipRGCs).
Melanopsin has a peak spectral sensitivity of approximately 480 nm, in the cyan part of the visible spectrum [16]. This action spectrum was first determined using melatonin suppression, another non-visual response to light [17, 18]. The spectral sensitivity curve did not match rods or any individual cone type which, along with work in blind and color-blind patients, strongly implicated the presence of a novel photopigment.
ipRGCs are sparse, comprising only about ~1% of all retinal ganglion cells, but are distributed across the entire retina in a network ideally suited for detecting overall ambient light level (irradiance) rather than high-resolution images [19]. They project via the RHT directly to the SCN, where they deliver the light signal that resets the clock daily. They also project to other brain areas involved in sleep, arousal and mood.
ipRGCs evolved to tell the brain whether it is day or night, essentially is the light on or off? They evolved earlier than the visual photoreceptors and are slower to be activated and deactivated than rods or cones, and do not respond to the rapid changes in light required for vision. They will even continue to send signals after the light stimulus has ended, which has implications for sleep as discussed later.
Note that the term ‘circadian’ photoreceptor is not entirely correct as the melanopsin-containing ipRGC mediate light inputs for a range of non-visual responses, not just entrainment of the circadian clock. The ability of light to suppress melatonin, the biochemical signal of darkness, to directly alert the brain, to elevate mood or constrict the pupils is also mediated primarily by the ipRGCs and does not involve resetting the circadian clock, so the term ‘non-visual’ photoreceptor is more broadly appropriate.
Finally, while melanopsin-containing ipRGCs are the primary photoreceptors for circadian entrainment and other non-visual responses, cone photoreceptors also contribute meaningfully to non-visual light responses, particularly in dim light or at the start of a light exposure [11, 20, 21].
The most compelling natural experiment illustrating the indispensability of light for circadian entrainment is the impact of total blindness on circadian entrainment. When the eyes are lost or when the entire retina is non-functional, the SCN receives no light information and the circadian clock becomes non-entrained, drifting each day according to its intrinsic period, typically slightly longer than 24 hours [8].
The resulting disorder is called Non-24-Hour Sleep-Wake Disorder (N24SWD), and it is experienced by the majority of totally blind people who have lost their eyes or whose retinae are entirely non-functional. The condition is sometimes, but not always, characterized by a cyclic pattern of sleep disturbance: the individual's sleep and wake times drift progressively later each day, so that for weeks at a time they may be trying to sleep during the day and lie awake at night, before the clock completes its slow rotation back into synchrony with the social day — at which point sleep temporarily improves before the cycle begins again [8].
The duration of this cycle is determined by the individual's intrinsic clock period. Someone whose clock runs 30 minutes per day longer than 24 hours — a period of 24.5 hours — will take 49 days to complete one full circadian cycle; someone with a period of 24.1 hours will take approximately 8 months. Neither the patient nor their physician may recognize the cyclic nature of the disorder; it may be mistaken for depression, insomnia, or narcolepsy depending on which phase of the cycle the patient presents in.
These studies highlight the importance of light and how we take its non-visual functions for granted. If the eyes are functional and you live in a 24-hour light-dark cycle, the clock automatically resets itself each day to keep you entrained. If you were unfortunate enough to lose your eyes today, you would start to exhibit non-24-hour rhythms tomorrow – the clock needs to be reset each and every day by the light -dark cycle. The only other way to reset the clock when light is absent is using daily administration of melatonin [22] or a melatonin agonist [23], which must be given at the same clock time every day to provide a substitute signal for the missing light.
Not all light is equal in its ability to reset the human circadian clock, suppress melatonin, or enhance alertness. Through decades of carefully controlled laboratory studies, scientists have characterized the key properties of a light stimulus that determine its biological impact including light timing, illuminance, duration, pattern, wavelength, and light history [24].
The single most important determinant of a light stimulus's circadian effect is when it occurs in the circadian cycle. Light can either shift the clock earlier (phase advance) or later (phase delay) based on the circadian time of the exposure.
This relationship is captured in the Phase Response Curve (PRC) for light. Under normal entrainment, light exposure from approximately 18:00 to 06:00 causes phase delays whereas light from approximately 06:00 to 18:00 causes phase advances. The maximum delaying effect occurs around 3–6 am, while the maximum advancing effect occurs around 6–9 am [25-27]. The PRC transitions through a ‘crossover point’ several hours before habitual wake time, and the precise timing of this crossover point differs between individuals depending on chronotype, termed the phase angle of entrainment [28-30].
This is why timing the light correctly to alleviate jet lag is crucial. Flying eastward requires phase advances where adapting to a westward shift requires a phase delay. Getting light exposure at the wrong time — as is often advised by well-meaning but non-expert sources — can shift the clock in the wrong direction and worsen jet lag. Similar problems can happen for shift workers if they see light at the wrong time, making it harder to adapt.
The circadian system responds to light illuminance level in a markedly non-linear fashion. Light levels typically experienced indoors can substantially suppress melatonin and shift the circadian clock when the exposure is sustained over several hours, with only 100 lux, far below the level of daylight, accounting for 50% of the maximum response [31, 32].
The dose-response relationship for illuminance reaches saturation at only several thousand lux, again far below the levels experienced outside at dawn or dusk, ~10,000 lux, with the most rapid changes in response occurring at typical room light levels (50-400 lux) [32, 33]. This means that moderate indoor lighting can exert meaningful circadian effects, and practical advice for circadian resetting must therefore consider indoor light as a powerful biologically active stimulus.
Given that melanopsin is maximally sensitive to light of approximately 480 nm (short-wavelength "blue" light), it follows that blue, or blue-enriched, "cool white" light sources, are more effective for circadian resetting, melatonin suppression and alerting the brain than red-shifted "warm white" sources at the same irradiance [20, 21, 34, 35].
In practical terms, the correlated color temperature (CCT) of a light source provides a rough proxy for its circadian potency: ‘cool’-looking light sources with a high CCT (5,000–6,500 K), which on average contain more short wavelength light, are more effective non-visual stimuli, while low CCT sources (2,700–3,000 K), which appear warm and orange-tinged, are less potent [36]. The appropriate scientific metric to quantify light for non-visual responses is to calculate the mel-EDI (melanopic equivalent daylight illuminance), which quantifies the light level as detected by the melanopsin system and is standardized in the CIE S 026 framework [37]. Using traditional photopic lux is not appropriate, as the lux sensitivity curve assumes that of the three-cone visual color vision system, which is inconsistent with the melanopsin-containing photoreceptors mediating the response.
The spectral composition of light is particularly important when considering the impact of electronic screens, such as smartphones, tablets, laptops, and television, as they emit substantial short-wavelength light. Evening use of these devices suppresses melatonin, delays the circadian clock, and impairs subsequent sleep quality [38, 39].
Longer light exposures produce larger circadian phase shifts, but the relationship is non-linear: shorter light exposures produce disproportionately large effects relative to subsequent hours [40]. For example, a 1-hour bright light exposure produces about a third of the phase shift achievable with a 6.5-hour exposure, despite only 15% of the duration, making shorter exposures relatively more efficient per minute of light [26].
Equally striking is the efficacy of intermittent light pulses. Gronfier and colleagues showed that six 15-minute pulses of bright light separated by 60-minute dim intervals (representing only 25% of the total time) produced approximately 75% of the circadian phase shift achieved by continuous 6-hour exposure [41, 42]. Even shorter pulses — as brief as 15 seconds of bright light — produce measurable circadian shifts [43]. The circadian system can therefore integrate light information across time, treating intermittent daytime light as a signal of "day" even when gaps exist between exposures. This approach has been used to provide practical countermeasures to improve alertness and performance during night shifts where continues exposures is not possible, for example in an ICU [44] or NASA Mission Control [45]
The findings also come with an important warning: brief unintended light pulses at night — getting up to use the bathroom, briefly checking a phone — can produce meaningful circadian phase shifts or alertness if they occur at the sensitive phase of the PRC or at night.
The circadian system does not respond to each light exposure in isolation; prior light history can modulate sensitivity to subsequent exposures. This phenomenon means that individuals who have been exposed to dim light earlier in the day show enhanced melatonin suppression or phase resetting responses to light later that night compared to individuals who spent the day in brighter dim light [46-48]. Desensitization can also occur, reducing sensitivity to subsequent light due to prior bright light exposure. These effects may also be triggered by changes in spectra not just illuminance level. The impact of these changes is not enormous and the exact mechanisms and characteristics of the light history effects are not fully understood but it is likely important to be aware of the sequence of light exposure changes when designing countermeasures rather than just consider the current conditions in isolation.
There is a subtle but important reciprocal relationship between sleep and light exposure: sleep itself gates the amount of light that reaches the circadian system. When we are asleep, our eyes are closed, and we are typically in a dark room, blocking light to the brain and providing a dark signal to the clock at the appropriate circadian time (night). Even if a light is on, the eyelids block about 95% of the light, and shift it to the red, and less effective, end of the spectrum as the light passes though skin and blood vessels. Conversely, when we are awake, our eyes are open and we are typically exposed to light. Even if this light is dim, it is seen by the brain as ‘light’ compared to when we close our eyes. The sleep-wake pattern therefore largely determines the relative pattern of the light-dark cycle that reaches the SCN each day, even if the environmental light is not that different [28, 49]. For example, a 10 lux light exposure might be interpreted as a daytime signal, and enough to entrain the circadian clock, if sleep occurs in darkness [50].
This relative light sensitivity of the circadian systems means that the timing of both light AND dark is crucial. If sleep occurs at the "wrong" circadian time, for example during the day in a shift worker, or when trying to sleep in a new time zone, brings with it a displaced pattern of light and dark exposure, which can then further shift the clock in the wrong direction. The result is a vicious cycle: circadian misalignment causes poor sleep at the desired time, and poor sleep at an inappropriate time worsens circadian misalignment. This mechanism underlies both jet lag and shift work disorder, and contributes to other circadian phase angle disorders.
Jet lag occurs when rapid transmeridian travel changes the light-dark cycle more rapidly than the circadian system can adapt. The symptoms, including problems sleeping at the new local night, excessive sleepiness during the new local day, impaired cognition, gastrointestinal disturbance, and mood changes, persist until the circadian system re-entrains to the new local time, which under unaided conditions typically takes at least one day per time zone crossed.
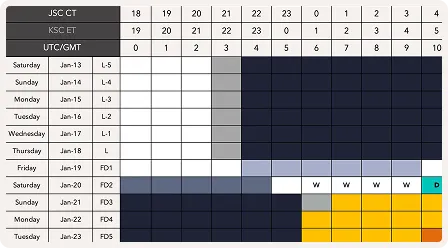
Strategically timed light and light avoidance, underpinned by application of the light PRC, is absolutely essential to any circadian readaptation intervention and can accelerate adaptation by three to four times compared to the unaided rate [25-27]. The Timeshifter jet lag app translates this complex science into personalized recommendations accounting for the individual's chronotype, direction of travel, itinerary, and sleep pattern.
Timeshifter's jet lag app is the most downloaded and highest-rated jet lag app in the world. Get personalized jet lag plans based on your sleep pattern, chronotype, itinerary, and personal preferences.
Learn more

Shift work disorder is similar to jet lag but is a more chronic form of circadian disruption during which the light-dark cycle also changes to rapidly for the circadian clock to keep up with due to changing work schedules. Night shift workers are required to be awake and see light during the night, at a time when their circadian clock is promoting sleep, and to sleep and see dark during the day when the clock promotes alertness. Approximately 20% of the work force experiences shift work, with another 20% altering their light-dark cycle by being ‘early-risers’, having to get up and go to work earlier than they naturally would, including a circadian phase angle disorder. The chronic circadian misalignment associated with shift work has been associated with increased risks of obesity, type 2 diabetes, cardiovascular disease, and certain cancers
Download the free shift work guide to alertness and safety.
Timeshifter's shift work app is an entirely new way for shift workers to optimize their sleep, alertness, and quality of life. Import your work schedule to get highly personalized advice.
Learn more
Beyond jet lag and shift work, misalignment between the timing of the internal clock and the desired sleep schedule underlies the circadian rhythm sleep-wake disorders (CRSWDs), including Delayed Sleep Wake Phase Disorder (DSWPD), in which the clock is set abnormally late, making early rising difficult; and Advanced Sleep Wake Phase Disorder (ASWPD), in which the clock is too early, causing excessive evening sleepiness and very early morning waking [51]. Even if the threshold to reach a clinical disorder is not reached, misalignment of wake and sleep cycles, and therefore light-dark cycles, is common in day-to-day life, for example when staying up late at the weekend, or having to wake up early for work, sometimes termed ‘social jet lag’. Treatment of these disorders relies primarily on strategically timed light exposure to shift the clock toward the desired phase [51].
Light does not only affect the circadian clock through entrainment; it is also a direct stimulant and acutely elevates alertness and arousal during the day and night, operating through mechanisms that are partially independent of circadian phase shifting [34-36, 52]. Brain imaging studies have shown that light exposure activates multiple regions implicated in alertness, attention, and cognition, including the thalamus, hippocampus, amygdala, and brainstem [12, 53].
This direct alerting effect of light has a secondary consequence for circadian regulation. If evening light increases alertness, for example by using electronic screens in the evening, it delays the onset of sleep — moving sleep (and therefore dark) to a later time. A later sleep onset means that the individual is awake (and thus exposed to light) during a circadian phase when light causes delay shifts, further postponing the clock. Conversely, waking up too early and seeing morning light simultaneously advances the clock directly and enhances alertness, shifting the clock to an earlier time. The change in light timing has a knock-on effect the next day, making it harder to fall asleep at normal time in the first example, and harder to sleep in in the second example, further reinforcing the circadian misalignment in a continuous feedback loop.
Light can therefore influence circadian rhythms through two pathways: a direct neural pathway via the RHT to the SCN, and an indirect pathway via light-induced changes in alertness and sleep timing that then alter the pattern of light-dark exposure received by the clock. Both pathways are important, and solutions to alleviate jet lag, shift work and circadian misalignment has to time light exposure appropriately taking both functions into account.
The Industrial Revolution, and particularly the invention of electric lighting in the late 19th century, gave humanity the ability to illuminate the night on a massive scale. In the United States alone, approximately 300,000 electric lamps were sold in 1885; by 1945, this had grown to 800 million annually [54]. Today, approximately a quarter of all electricity generated globally is used for lighting. This transformation of the nocturnal environment has proceeded at a pace far exceeding the capacity of evolutionary adaptation.
Man-made light at night now permeates most human living environments, entering bedrooms from street lighting, advertising, and neighboring buildings. This light can disrupt the regular daily light-dark cycle exposure that the circadian system craves, blurring the biological signals of night and day.
Epidemiological studies have shown associations between residential exposure to outdoor light at night and increased risk of breast cancer, obesity, and depression, though causality has not been definitively established and confounding is difficult to exclude in observational research. Shift workers and healthcare workers — populations with the highest chronic exposure to light at night — show the most consistent evidence of adverse health effects [55, 56].
Beyond humans, the ecological consequences of artificial light at night are extensive and better established. Light pollution disrupts the breeding, foraging, migration, and predator-prey dynamics of a vast range of species [57]. Nocturnal insects — critically important as pollinators and as food for birds and bats — are attracted to and disoriented by man-made light, leading to population declines [58]. Migrating birds navigate by stars and moonlight and are confused by urban skyglow, leading to fatal collisions with illuminated buildings [59], and light at night disrupts behavior and reproduction [60]. Sea turtle hatchlings follow the moonlight toward the ocean but are drawn inland by beachfront lighting [61].
A concerning recent development is the proposed plan to use mirrors orbiting the Earth to create areas of bright light anywhere on Earth at any time of day or night, potentially disrupting the daily light-dark cycles nearly all organisms on the planet rely on to time their biology [62]. These effects cascade through ecosystems in ways that are only beginning to be understood and such massive changes to the natural light-dark cycle should be resisted.
Practical solutions to current light pollution are available and relatively simple: directing luminaires downward (not upward or horizontally), using motion sensors and timers, selecting spectrally appropriate light sources, and simply switching off lights when not needed. These measures simultaneously reduce energy consumption, protect human health, and benefit other species — a rare convergence of individual, societal, and ecological interests.
Light is far more than a sensory stimulus enabling us to see. It is the primary timekeeper of the human body, synchronizing a near-24-hour internal clock that orchestrates sleep, hormones, metabolism, cognitive performance, immune function, and disease risk. This timekeeping function is primarily mediated by a dedicated photoreceptor system, the melanopsin-containing ipRGCs, that projects directly to the central circadian clock in the SCN.
The consequences of getting light wrong are not trivial: Totally blind individuals unable to entrain to the 24-hour day suffer N24SWD; shift workers face increased chronic health problems and higher rates of accidents and injuries; and urban populations experience challenging light-dark cycles through electronic device use and man-made light at night. The technology that enables our modern 24/7 society has long-term health consequences that we are only just realizing. Light is something most of us take entirely for granted — until, like the blind, or a shift worker, it is taken from us, misaligned, or cycling at the wrong frequency.
As we illuminate our cities ever more brightly and carry glowing screens into our bedrooms each night, understanding how light shapes our biology has never been more important. Protecting the natural light-dark cycle — for ourselves and for the other species who share our illuminated world — is one of the simplest and most powerful things we can do for human and planetary health.